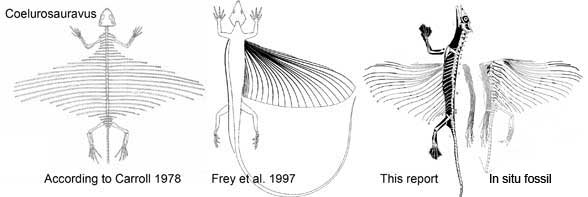
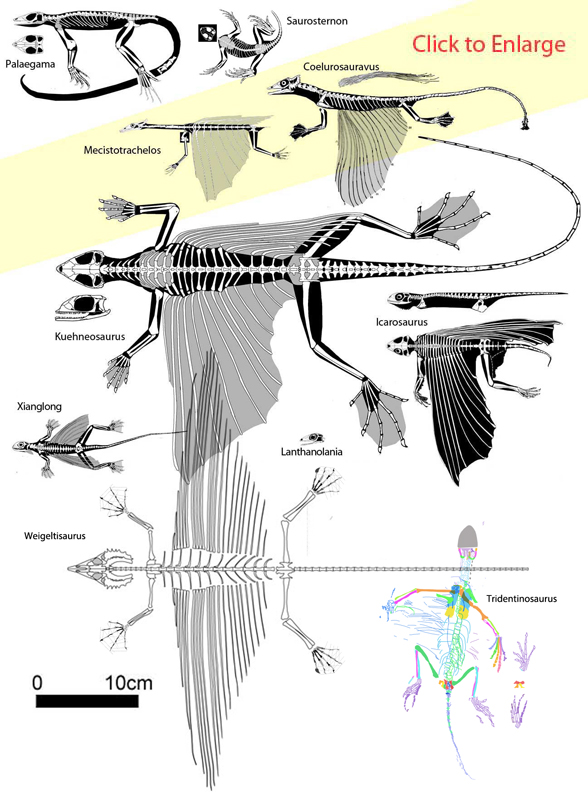
Carroll RL 1978. Permo-Triassic "Lizards" from the Karoo System Part II. A gliding reptile from the Upper Permian of Madagascar. Palaeontografica Africana. 21:143-159.
Dal Piaz Gb. 1932 (1931). Scoperta degli avanzi di un rettile (lacertide) nei tufi compresi entro i porfidi quarziferi permiani del Trentino. Atti Soc. Ital. Progr. Scienze, XX Riunione, v. 2, pp. 280-281. [The discovery of the remains of a reptile (lacertide) in tuffs including within the Permian quartz porphyry of Trentino.]
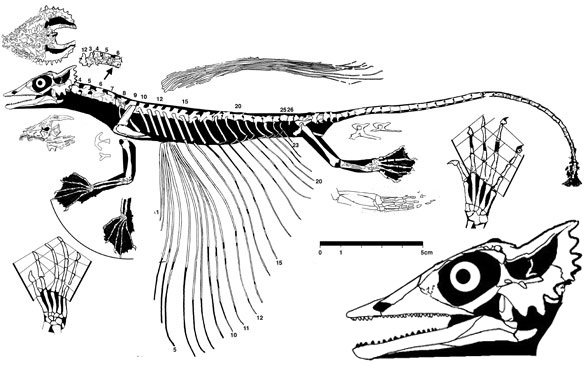
Evans SE 1982. Gliding reptiles of the Late Permian. Zoological Journal of the Linnean Society, 76:97–123.
Evans SE and Haubold H 1987. A review of the Upper Permian genera Coelurosauravus, Weigeltisaurus and Gracilisaurus (Reptilia: Diapsida). Zoological Journal of the Linnean Society 90:275–303.
Fraser NC, Olsen PE, Dooley AC Jr and Ryan TR 2007. A new gliding tetrapod (Diapsida: ?Archosauromorpha) from the Upper Triassic (Carnian) of Virginia. Journal of Vertebrate Paleontology 27 (2): 261–265.
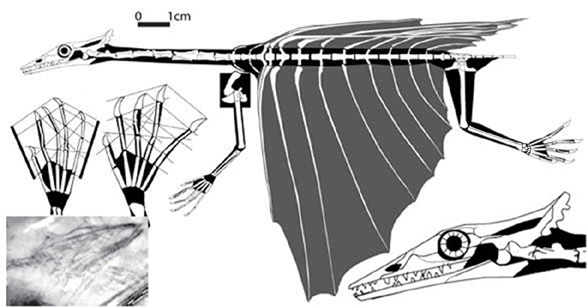
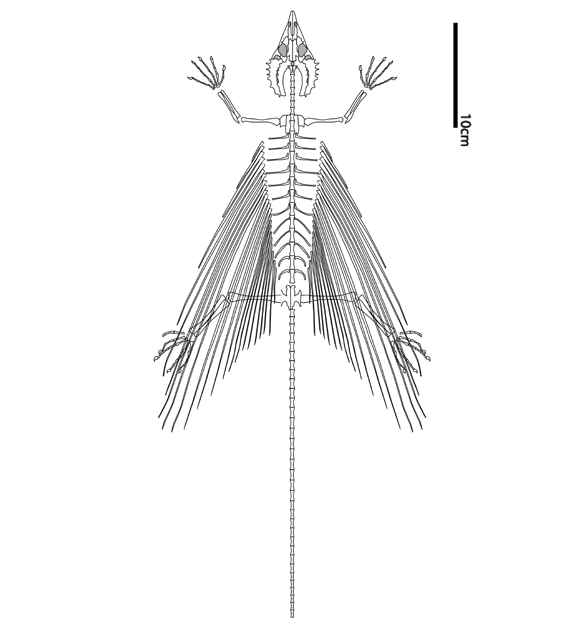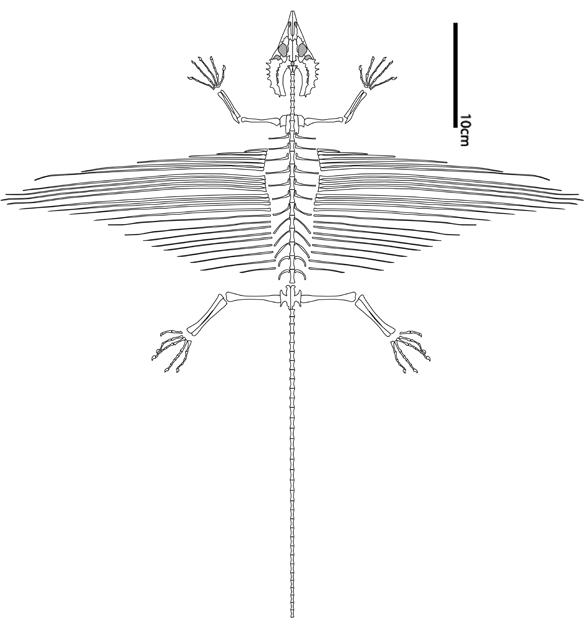
Frey E, Sues H-D and Munk W 1997. Gliding Mechanism in the Late Permian Reptile Coelurosauravus. Science Vol. 275. no. 5305, pp. 1450 - 1452
DOI: 10.1126/science.275.5305.1450
Leonardi P 1959. Tridentinosaurus antiquus Gb. Dal Piaz, rettile protorosauro permiano del Trentino orientale. Memorie di Scienze Geologiche 21: 3–15.
Piveteau J 1926. Paleontologie de Madagascar, XIII. Amphibiens et reptiles permiens: Annales de Paleontologie, v. 15, p. 53-128.
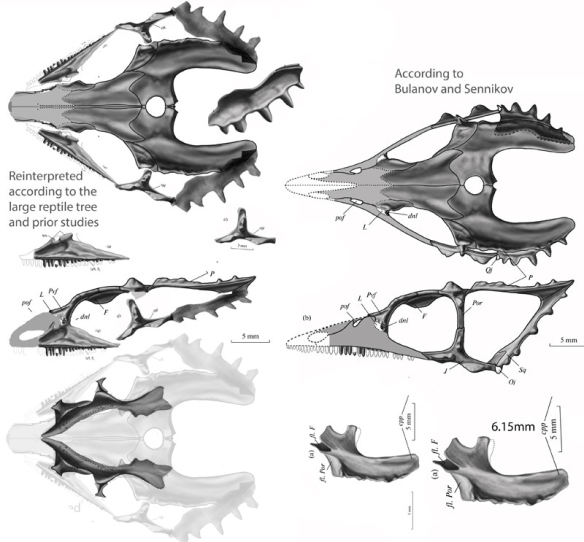
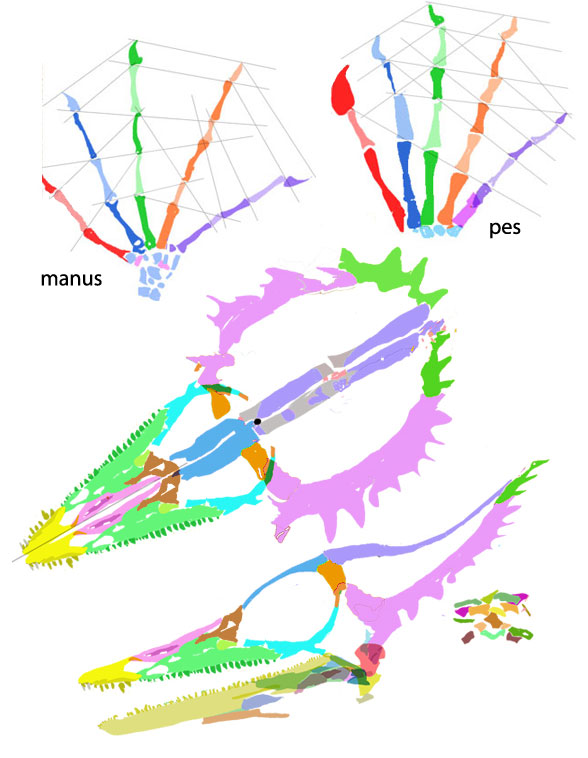
Pritchard AC, Sues H-D, Scott D, Reisz RR. 2021. Osteology, relationships and functional morphology of Weigeltisaurus jaekeli (Diapsida, Weigeltisauridae) based on a complete skeleton from the Upper Permian Kupferschiefer of Germany. PeerJ 9:e11413 DOI 10.7717/peerj.11413
Weigelt J 1930. Palaeochameleo jaekeli nov. gen., nov. sp., ein neuer Rhynchocephale aus dem
Mansfelder Kupferschiefer. Leopoldina 6:625–642.
wiki/Coelurosauravus
wiki/Mecistotrachelos
wiki/Tridentinosaurus - not yet created
wiki/Weigeltisaurus |